

הפיתול המצחי התחתון
 |
הפיתול המצחי התחתון (באנגלית: (inferior frontal gyrus (IFG), הוא פיתול בקליפת המוח, התחתון מבין ארבעת הפיתולים שבאונה המצחית וחלק מן האונה הקדם מצחית. הפיתול תחום מלעיל על ידי החריץ המצחי התחתון (אנ') המפריד אותו מן הפיתול המצחי האמצעי (אנ'). גבולו התחתון הוא החריץ הצידי (אנ') אשר מפריד אותו מהפיתול הרקתי העליון (אנ') והגבול האחורי לפיתול המצחי התחתון הוא החריץ הקדם מרכזי (אנ') המפריד אותו מהפיתול הקדם-מרכזי[1].
כמו כן, הפיתול המצחי התחתון מכיל את אזור ברוקה, אזור המעורב בעיבוד לשוני, ובהפקת שפה.
מבנה

נהוג לחלק את הפיתול המצחי התחתון לשלושה אזורים נפרדים[2][3][4]: החלק המכסי (pars opercularis - Pop), החלק המשולשי (pars triangularis - PT) והחלק הארובתי (the pars orbitalis - POr). חלוקה זו נקבעה בעקבות חריצים קטנים היוצאים מן החריץ הצידי הנקראים ענפי (Ramus) החריץ הצידי[5]. החלק המכסי של הפיתול המצחי התחתון הוא חלק מן קליפת המוח המצחית המכסה את האינסולה ומכאן שמו. החלק המשולשי הוא בעל צורת משולש ואילו החלק הארובתי נמצא קרוב לאזור ארובת העין והוא חלק מן קליפת המוח הארובתית-מצחית. שלושת החלקים מקושרים ביניהם רבות. דבר זה נמצא במחקר אשר ביצע גרייה חשמלית בחלקים נפרדים בפיתול אצל בני אדם תחת הליך נוירוכירוגי, וראה כי ההיטל של הגרייה מגיע לכל אורך הפיתול[6] עובדה זו נתמכה גם על ידי מחקרים נוספים[7][8].
בחלוקה לאזורי ברודמן החלק המכסי של הפיתול המצחי התחתון ידוע כאזור ברודמן 44, החלק המשולשי של הפיתול המצחי התחתון ידוע כאזור ברודמן 45 ושניהם יחדיו מהווים את אזור ברוקה. כמו כן החלק הארובתי של הפיתול המצחי התחתון ידוע כאזור ברודמן 47.
פעילות

ידוע עוד מהמאה ה-19, על ידי ניסויי הלזיות של ברוקה[9], כי ה-IFG מעורב ביכולות שפתיות. תיקופים נרחבים המתפרסים על מתודות שונות נצברו עם השנים[10][11][12], והראו כי האזור מעורב במספר רב של פעילויות שפתיות הכוללות אף זיהוי מחוות רגשיות בדיבור[13]. מתוך הפעילויות השפתיות ה-Pop שבהמיספרה השמאלית מקושר לקליפת המוח הקדם-מוטורית ולאינסולה הקדמית ברשת עצבית המפיקה את ההברות הבסיסיות, והאזור POr ידוע כקשור לעיבוד מבנים מוזיקליים[14]. בכללו, מניסויים שהשמשו במכשיר הפוגע זמנית בתפקוד האזור (TMS), התגלה כי הפיתול מפחית למידה של מידע המזוהה כלא נעים. זאת משום שהשימוש ב-TMS החיש את יכולת הלמידה של מידע שכזה. נוסף על כך פעילות מוגברת של ה-IFG נמצאת בהתאמה להימנעות מלקיחת סיכונים[15]. פעולה נוספת שידוע כי האזור מעורב בה, לפחות חלקית, הוא הפעלת ה-(mismatch negativity (MMN (אנ'). ה-MMN הוא הופעת דפוס פוטנציאלים חשמליים ייחודי על הקרקפת בתגובה לגירוי שסותר את הזיכרון מהשניות שקדמו לגירוי. אפשרות נוספת לפרשנות ה-MMN היא שהוא בא כתגובה למידע הסותר את הניבוי של המערכת הקוגניטיבית על הסדר של הסביבה החיצונית[16]. לדוגמה משפט עם שגיאות תחביר בין שלל משפטים תקינים יעורר MMN. ל-MMN ידוע אזור אשר גורם לתגובה הנמצא על הפיתול הרקתי העליון, אך עם זאת' מחקרים רבים מצאו מקור נוסף הנמצא באזורים מצחיים. במקרים רבים באמצעות ניתוח מקורות של פריסת הפוטנציאלים ממוקר האזור המצחי על ה-IFG[17].
אזור ברוקה
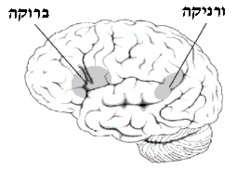
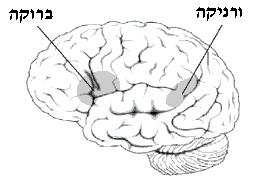
תרשים של צידו השמאלי של המוח המציג את אזור ברוקה ואזור ורניקה
 ערך מורחב – אזור ברוקה
ערך מורחב – אזור ברוקה
האזורים PT יחד עם אזור Pop מהווים את אזור ברוקה, אזור בסיסי בהפקת שפה. בגיעה באזור זה תגרום באופן ישיר לאפזיית ברוקה, אפזיה המאופיינת לנזק בלתי הפיך ליכולת הדיבור השוטפת והכתיבה, ומכונה אפאזיה אקספרסיבית(אנ'). פגיעה זו מתאפיינת ב"דיבור טלגרפי", שימוש חוזר במילים בודדות וקושי ביצירת משפטים, זאת למרות הבנה מלאה של השפה המדוברת או הכתובה.
נוירוני מראה
- ערך מורחב – נוירון מראה
נוירוני מראה הינם נוירונים הפעילים לא רק בעת ביצוע פעולה מוטורית אלא גם בעת התבוננות בעת אדם אחר העושה את אותה הפעולה. ההשערה הרווחת במחקר היא שפעילות הנוירונים מאפשרת למידה על ידי חיקוי ויכולת להבין את האחר, כאשר האחרונה עומדת בבסיס תהליכי תיאוריה של תודעה. יחד עם מקור באונה הפריאטלית, אחד המקורות המרכזיים ביותר של נוירונים אלו נמצע על החלק האחורי של ה-IFG הקרוב לקורטקס המוטורי[18].
קישורים חיצוניים
הערות שוליים
- ↑ Nolte (2002), The Human Brain, ISBN 978-0-323-01320-8 photos on p526 & p.546
- ↑ Brodmann K. 1909. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig: Barth.
- ↑ Economo C, Koskinas GN. 1925. Die Cytoarchitektonik der Hirnrinde des erwachsenen Menschen. Vienna/Berlin: Springer.
- ↑ Petrides M, Pandya DN. 2004. The frontal cortex. In: Mai JK, editor. The human nervous system. San Diego, CA: Elsevier. p 951–974.
- ↑ החריצים העליונים של חריץ קליפת המוח הצידי, אתר המילון הרפואי
- ↑ Jeremy D.W. Greenlee; Hiroyuki Oya; Hiroto Kawasaki; Igor O. Volkov; Meryl A. Severson III; Matthew A. Howard III; John F. Brugge (2007). "Functional connections within the human inferior frontal gyrus". The Journal of Comparative Neurology. 503 (4): 550–559. doi:10.1002/cne.21405. PMID 17534935.
- ↑ Henry RG, Berman JI, Nagarajan SS, Mukherjee P, Berger MS. 2004. Subcortical pathways serving cortical language sites: initial experience with diffusion tensor imaging fiber tracking combined with intraoperative language mapping. Neuroimage 21: 616–622.
- ↑ Petrides M, Pandya DN. 2002. Comparative cytoarchitectonic analysis of the human and the macaque ventrolateral prefrontal cortex and corticocortical connection patterns in the monkey. Eur J Neurosci 16: 291–310.
- ↑ ראה תרגום לאנגלית של מאמר המקור - von Bonin G. 1950. Remarks on the seat of the faculty of articulate language, followed by an observation of aphemia. In: Essay on the cerebral cortex. Springfield, IL: Charles C. Thomas. p 49–72.
- ↑ Lazar RM, Marshall RS, Pile‐Spellman J, Duong HC, Mohr JP, Young WL, Solomon RL, Perera GM, DeLaPaz RL. 2000. Interhemispheric transfer of language in patients with left frontal cerebral arteriovenous malformation. Neuropsychologia 38: 1325–1332.
- ↑ Dhond RP, Buckner RL, Dale AM, Marinkovic K, Halgren E. 2001. Spatiotemporal maps of brain activity underlying word generation and their modification during repetition priming. J Neurosci 21: 3564–3571.
- ↑ Otsuki M, Soma Y, Koyama A, Yoshimura N, Furukawa H, Tsuji S. 1998. Transcortical sensory aphasia following left frontal infarction. J Neurol 245: 69–76.
- ↑ Gernsbacher MA, Kaschak MP. 2003. Neuroimaging studies of language production and comprehension. Annu Rev Psychol 54: 91–114.
- ↑ Levitin, D. J., & Menon, V. (2003). Musical structure is processed in “language” areas of the brain: a possible role for Brodmann Area 47 in temporal coherence. Neuroimage, 20(4), 2142-2152.
- ↑ Christopoulos, GI.; Tobler, PN.; Bossaerts, P.; Dolan, RJ.; Schultz, W. (Oct 2009). "Neural correlates of value, risk, and risk aversion contributing to decision making under risk". J Neurosci. 29 (40): 12574–83. doi:10.1523/JNEUROSCI.2614-09.2009. PMC 2794196. PMID 19812332.
- ↑ Winkler, I. (2007). Interpreting the mismatch negativity. Journal of Psychophysiology, 21(3-4), 147-163.
- ↑ Deouell, L. Y. (2007). The frontal generator of the mismatch negativity revisited. Journal of Psychophysiology, 21(3-4), 188-203.
- ↑ Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annu. Rev. Neurosci., 27, 169-192.

שגיאות פרמטריות בתבנית:מיון ויקיפדיה
שימוש בפרמטרים מיושנים [ דרגה ] הפיתול המצחי התחתון26769732